Science
Related: About this forumInterrogating the Development of Ant Castes with Interfering RNA.
The paper in the primary scientific literature I'll discuss in this post is this one: Social regulation of a rudimentary organ generates complex worker-caste systems in ants (Ehab Abouheif et al Nature Volume 562, pages 574–577 (2018))
I have a decent understanding of the molecular biology of proteins, including the remarkable nuclear proteins that choreograph the behavior the behavior of DNA, the highly conserved - because they're optimized across species - four histones of chromatin. Lipid chemistry is highly complex and important, but it's fairly easy to wrap one's head around it without too much effort; I've had to think about lipids a lot in my life time and seldom feel intimidated when I have to do so. Sugars, because of their complex stereochemistry and their large arrays of functional groups are often beyond me, but to the extent I have been involved with them, it's all been pretty much about N,O glycosylation of proteins, which sometimes plays a huge role in proteomics, but, well, I can generally get by by looking at the pictures and the schematics. I actually don't know too many experts in sugar chemistry because there aren't all that many synthetic sugar drugs, although there are indeed sugar based drugs on the market: In fact, one of the oldest drugs commercialized, digoxin, is technically a glycolipid. The absence of sugar drugs is probably a function of the difficulty of sugar chemistry. I'm always amused by this excerpt from a wonderful text on sugar chemistry, especially the lines I have bolded here: The Sugar Code that reads:
In other words, people shy away from sugar chemistry because sugar chemistry is hard.
This text also suggests that the biochemistry of the nucleic acids is better understood because it's easier.
Well perhaps...
There are, now, a few drugs on the market that are in effect nucleic acids or modified nucleic acids, in particular anti-sense compounds. Also a major gene insertion product has been approved, Luxturna which cures a certain type of blindness with a single dose. By the way drugs which cure diseases have more economic difficulty than maintenance drugs like blood pressure pills and statins which only address symptoms or prevent - so long as they are continually used - disease. Luxturna represents an interesting ethical case, because I believe that a single dose - the only dose ever needed - to the eye costs close to a million dollars.
Anyone who has worked in a lab in a research capacity knows about the long path of training and education, long hours, disappointment after the acquisition of expensive and rare materials, the danger, the cost. I can only imagine the work that went into making Luxturna into a marketed product - the cost - the fear that the product would fail, the terror at looking at results and traveling to regulatory meetings. If one knows this, one finds that scientists deserve to be paid, but on the other hand, if someone is blind and can be cured of blindness, one also feels that it should be done for free.
(I don't have an answer for this conundrum, but I do know that tax breaks for shitheads like the orange President and his family at the expense of the rest of the world's population is criminal.)
Anyway...
On a superficial level, there is a beautiful simplicity to nucleic acids, with only five core building blocks in contrast to the 20 core building blocks of proteins (if one ignores post-translational modifications like the arginine -> ornithine or citrulline, etc, etc and bacterial codons for things like selenomethionine, phosphorylation, etc, etc and the aforementioned glycosylations.)
However, this superficial understanding conveys very little of the complexity of nucleic acid biochemistry; one of the great discoveries of recent times and applicable to the nature/nuture issue (or perhaps the "exposiome" ) is the marvelous complexity revealed in epigenetics.
This said, I have a very superficial working knowledge of these compounds and some of the experimental notes in the paper cited at the outset on ants inspires me to learn more - since there are parts of the nuts and bolts that I'll excerpt here that are clearly beyond me.
I don't know about you, but I find social insects, as problematic as some species are - yellow jackets and carpenter ants come to mind - fascinating. Before I knew any science whatsoever beyond the very primitive concepts discussed in elementary school in my time, I recall spending hours upon hours just watching sugar ants; sometimes feeding them just to watch them work.
The abstract of the nature paper refers to this wonder:
The authors explore the developmental molecular biology of the development of soldier ants pretty much from the very same genome of minor worker ants and for that matter, winged ants, queens and drones.
Soldier ants have big heads - and even if General Bonespurs in the White House is a long standing coward with an absurdly big head and thus is no soldier - in soldier ants big heads and powerful jaws are important in defending the colony from attack. The authors note that the soldier ants also have vestigial wings that minor worker ants lack and they explore the origins of this anatomical feature and its relation to big heads.
They do this by shutting off the expression of some genes by using interfering RNA on the messenger RNA produced by the DNA of the organism at different developmental phases. Interfering RNA is RNA that is complementary to expressed RNA and thus can interact with it forming double stranded (ds) RNA - as opposed to the functional single stranded RNA - thus blocking its ability to direct protein synthesis and ultimately function.
Some pictures from the text:
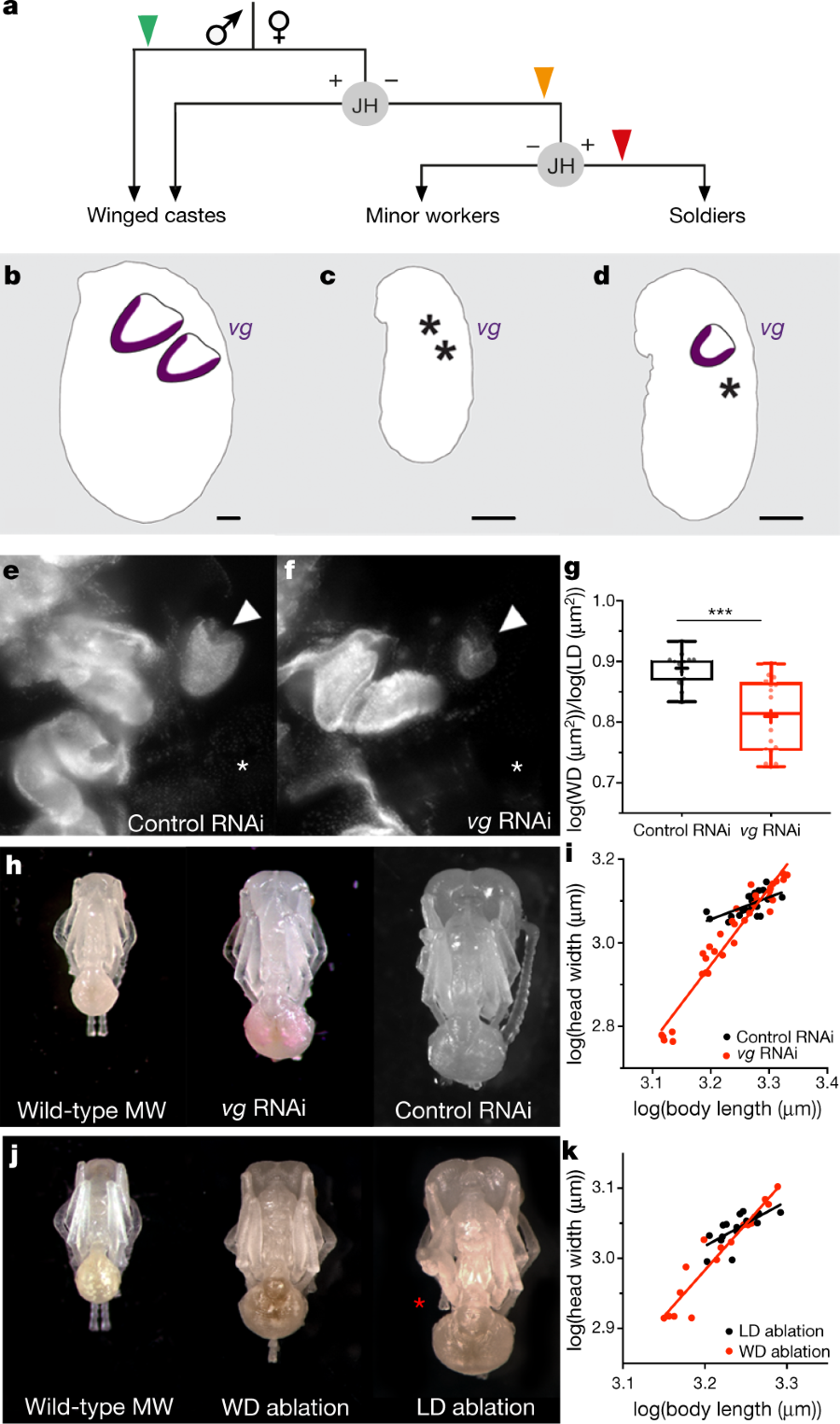
The caption:
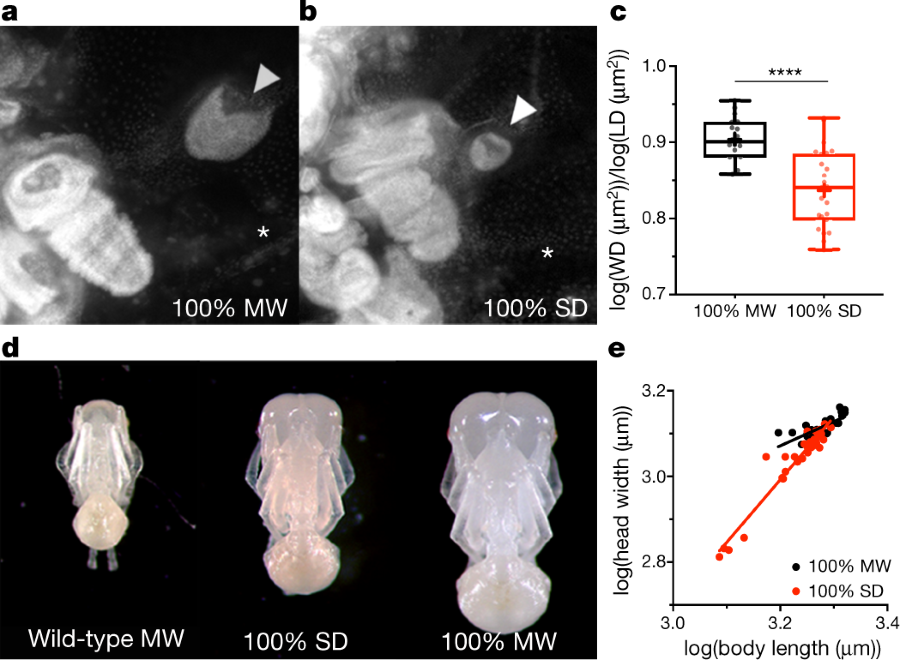
The caption:
A nice cartoon:
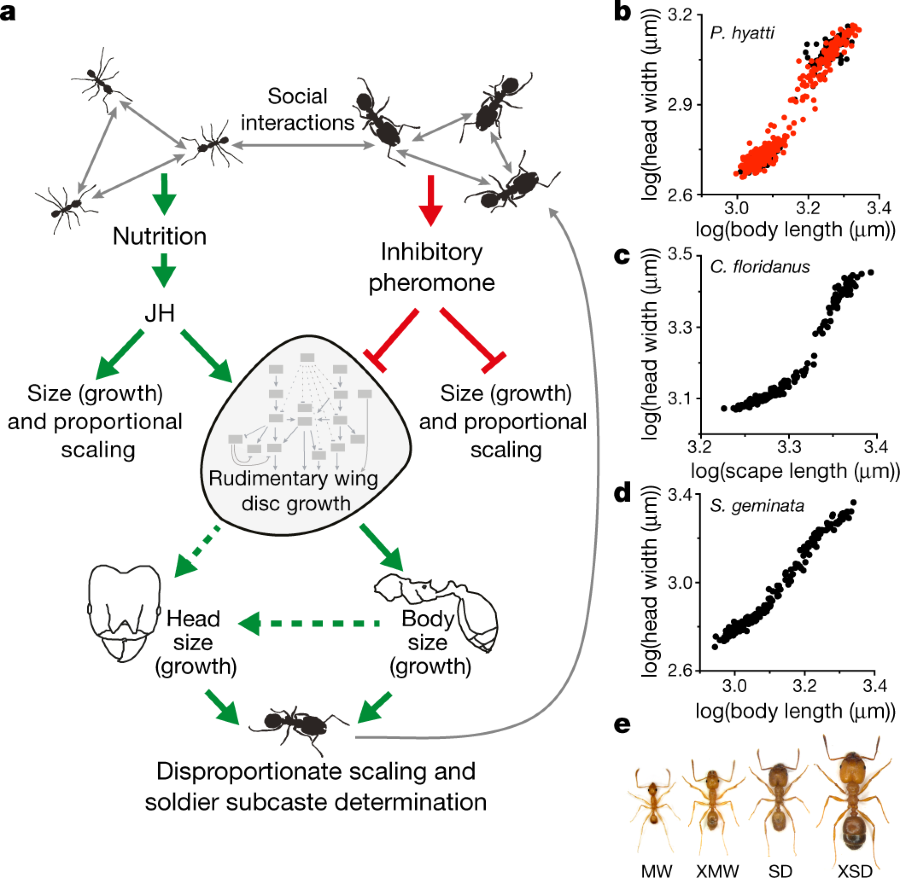
The caption:
And now some experimental text that is frankly over my head, but inspires me to learn more - I've ordered several texts from various libraries:
Forward: 5′-TATCCTTACCTKTAYCARACCC-3′ and reverse: 5′-GCTGACTATTCCAAAAGGARGG-3′ degenerate primers were designed based on a vestigial sequence alignment of Tribolium castaneum (XM_008201106), Apis mellifera (XM_001122002.4) and several ant species obtained from the Ant Genome Portal database (http://hymenopteragenome.org/ant_genomes/). P. hyatti RNA was isolated using TRIzol (Invitrogen) from a pool of embryos and larvae of different developmental stages. RNA was then reverse-transcribed to synthesize a cDNA library. PCR amplicons were ligated into the pGEM-T easy vector (Promega) and subsequently sequenced (MH683613) using Sanger sequencing at the Genome Quebec Innovation Centre at McGill University. To determine whether these vestigial sequences—including the TONDU/vestigial domain—are conserved, amino acid alignment was performed using Geneious Alignment on Geneious (R8), followed by manual alignment (Supplementary Fig. 2)...
...emi-quantitative reverse-transcription PCR of vestigial transcripts in P. hyatti
vestigial expression in wild-type terminal P. hyatti larvae was determined through the detection of vestigial transcripts using semi-quantitative PCR with reverse transcription (Extended Data Fig. 3 and Supplementary Fig. 1). The housekeeping gene Elongation factor 1 alpha (EF1a) was used as our reference gene because it has previously been validated as a reference gene for quantitative PCR with reverse transcription in social insects45. Therefore, EF1a was first cloned and sequenced (MH683615) from a P. hyatti cDNA library using the following degenerate primers: forward 5′-GATTCYGGCAAGTCGACCA-3′ and reverse 5′-GGAACTCTTGGAAAGCCTCAAC-3′. The PCR product was sequenced using Sanger sequencing at the Genome Quebec Innovation Centre at McGill University. To extract RNA, three minor worker larvae and three soldier larvae at the terminal stage were collected from a laboratory colony and total RNA was extracted from minor workers and soldiers separately. The tissue was disrupted using a TissueLyser (Qiagen) bead mill and RNA was extracted using the TRIzol (Invitrogen) RNA extraction protocol46, and then purified using the RNeasy Plus Kit (Qiagen). Minor worker and soldier RNA was treated with DNase I (Invitrogen) to remove genomic DNA before being reverse transcribed into cDNA using the Superscript III First-Strand Synthesis System (Invitrogen). The concentrations of total RNA and total cDNA were normalized between minor workers and soldiers before cDNA synthesis and PCR, respectively. The two cDNA libraries were used as PCR templates for the semi-quantitative PCR with reverse transcription of vestigial and EF1a. The P. hyatti vestigial PCR primers used were: forward 5′-TCCTTACCTGTATCAGACCCATC-3′ and reverse 5′-TGTCGATCTGTCGTCGTCCAA-3′, and the P. hyatti EF1a PCR primers used were: forward 5′-TCAGGACGTGTACAAGATC-3′ and reverse 5′-CAATGACCTGTGCAGTAAAG-3′. The PCR was performed using an annealing temperature of 56 °C with 31 thermocycles and four serial dilutions of the two cDNA libraries.
I'm an old man, but being old, and recognizing that there isn't much time left, I feel more and more urgency about finding things out.
Youth - and certainly my youth - as they say in a wonderful platitude that became a platitude by being true, is wasted on the young.
If nothing else, I've tried to impress that on my sons, and happily they both seem to get it, more or less.
A beautiful thing on a rainy Saturday.
Have a nice weekend.
